ДНК-вакцина

ДНК-вакцина (также генная вакцина, вакцина на основе нуклеиновых кислот) — генно-инженерная конструкция, которая после введения в клетку обеспечивает продуцирование белков патогенов или опухолевых антигенов и вызывает иммунную реакцию. Введение ДНК-вакцин в организм называют генетической иммунизацией. ДНК-вакцинация имеет ряд преимуществ по сравнению с обычными вакцинами. В частности, показано, что такие вакцины обеспечивают не только выработку антител (гуморальный иммунитет), но и специфический цитотоксичный ответ (клеточный иммунитет), что ранее было достижимо только с помощью живых вакцин. Сегодня ДНК-вакцины не применяют для лечения человека, однако прогнозируется, что генетическая иммунизация поможет преодолеть целый ряд заболеваний.
История создания[править | править код]
Идея использовать фрагменты ДНК для вакцинации появилась в 50-60-е годы. После серии опытов было установлено, что генетическая информация ДНК сохраняет способность транскрибироваться и транслироваться после переноса в другую клетку[1][2]. В том же году обнаружили, что введение животным генома вируса полиомиелита стимулирует выработку антител[3][4]. Позже активацию гуморального иммунитета показали для молекул ДНК, полученных из неинфекционных агентов[5]. Начиная с 90-х годов научные лаборатории начали всё активнее исследовать иммуностимулирующие свойства ДНК. В 1992 году Танг вместе с коллегами показал, что ген гормона роста человека, встроенный в плазмиду, стабильно экспрессируется в организме мыши, а синтезированный гормон распознаётся иммунной системой как антиген и стимулирует выработку антител. Процесс ввода плазмидной ДНК для стимуляции гуморального иммунитета был назван Тангом «генетическая иммунизация»[6]. Однако уже в следующем году другая группа учёных заявила, что введение плазмиды, кодирующей белки вируса гриппа, вызывает как гуморальный, так и клеточный ответ[7]. Индуцирование обеих ветвей иммунитета в том же году обнаружили и для плазмиды, содержавшей гены ВИЧ[8]. С 1995 года начали появляться данные, что ДНК-вакцинация способна активировать иммунную систему против раковых заболеваний[9][10]. Около 20 лет назад состоялись первые клинические испытания ДНК-вакцин, которые прежде всего должны были продемонстрировать безопасность нового метода. Пациентам вводили гены ВИЧ, вируса гриппа, герпеса, гепатита B, возбудителя малярии. Результаты всех тестов оказались вполне обнадеживающими: ДНК-вакцины стабильно экспрессировались, провоцировали иммунный ответ и не вызвали серьезных побочных эффектов, что стало толчком для их дальнейшего исследования[11].
Конструирование ДНК-вакцины[править | править код]

По структуре ДНК-вакцина — это встроенная в вектор нуклеотидная последовательность, кодирующая определённый антиген или антигены. Вектором в генной инженерии называют молекулу нуклеиновой кислоты, которая служит для доставки генетического материала в клетки и обеспечивает его репликацию или экспрессию. Ранее для транспортировки генов в клетку применяли векторы на основе вирусов: модифицированного (ослабленного) вируса натуральной оспы, аденовирусов и ретровирусов. Вирусные векторы являются достаточно эффективными, однако имеют значительную вероятность развития побочных эффектов, связанную с относительно высокой иммуногенностью самого вектора. Поэтому на сегодня в качестве вектора чаще используют бактериальную плазмиду — небольшую стабильную кольцевую молекулу ДНК, способную к автономной репликации. Сама по себе плазмида не вызывает нужного специфического иммунного ответа, для этого в неё вшивают гены иммуногенных белков. Также ДНК-вакцина должна содержать регуляторные последовательности, необходимые для экспрессии генов в клетках эукариот. Готовую ДНК-конструкцию доставляют в бактериальную клетку, где наращивается количество её копий. После этого проводят выделение и очистку плазмид, которые несут нужную вставку[12].
Дизайн плазмидного вектора[править | править код]
Важным этапом создания ДНК-вакцин является дизайн (конструирование) вектора. Обязательными структурами плазмидного вектора являются сайты рестрикции, селективный маркер и точка начала репликации ДНК-вакцины в бактериальной клетке. Чтобы осуществлялся синтез антигена, ДНК-вакцина должна содержать промотор и сигнал полиаденилирования. Промотор является важным фактором эффективности вакцины, поскольку определяет силу иммунного ответа: чем больше экспрессия гена, кодирующего вирусный или опухолевый антиген, тем сильнее иммунный ответ. Чаще всего используют промотор вируса SV40 или цитомегаловируса (CMV). Для стабилизации мРНК-транскриптов в плазмиду встраивают сигнал полиаденилирования, чаще всего полученный из гена гормона роста быка (BGH) или вируса SV40. В качестве селективных маркеров выбирают бактериальные гены устойчивости к антибиотикам; часто это ген устойчивости к канамицину. При конструировании ДНК-вакцин наиболее популярна точка начала репликации Escherichia coli[13].
Выбор гена для иммунизации[править | править код]
Вектор является важным компонентом ДНК-вакцины, однако её иммуногенность определяется именно вставкой — последовательностью ДНК, которая кодирует антиген. Среди вирусных антигенов для иммунизации лучше всего подходят белки слияния — это относительно консервативные белки, которые обеспечивают проникновение вируса в клетку. Для вакцинации против грамположительных бактерий в плазмидный вектор целесообразно встраивать гены тех бактериальных белков, которые определяют патогенез заболевания. Среди белков грамотрицательных бактерий высокую иммуногенность имеют порины[14]. Для терапевтических противоопухолевых ДНК-вакцин используют белки-маркеры раковых клеток[15].
Способы доставки ДНК-вакцин в клетку[править | править код]
Готовую вакцину нужно доставить в организм человека или животного, где её точка назначения — антигенпрезентирующие клетки (АПК) — макрофаги, дендритные клетки, B-лимфоциты. Здесь будет происходить синтез и посттрансляционная модификация антигена, после чего он будет встроен в мембрану клетки, чтобы привлечь внимание иммунной системы. Основная проблема заключается в доставке достаточного количества плазмиды в АПК. Методы доставки генетического материала в клетку обычно разделяют на 2 группы: вирусные и невирусные. Поскольку вирусные векторы имеют ряд существенных недостатков, в данном разделе представлены лишь невирусные методы доставки ДНК-вакцин.
- Схематическое изображение клеточных процессов при введении ДНК
-
 С помощью аденовируса
С помощью аденовируса -
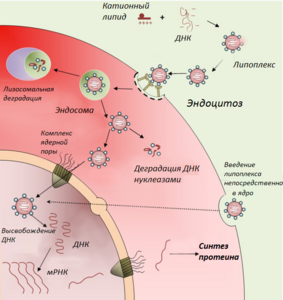 В составе липоплекса
В составе липоплекса


Микроинъекция[править | править код]
В начале 1990-х для введения ДНК в клетку наиболее распространёнными были внутримышечные микроинъекции, что обусловлено простотой метода. Для этого ДНК растворяют в воде или изотоническом растворе, при необходимости добавляют адъювант (вещество, которое усиливает иммунный ответ). Далее с помощью тонкой стеклянной трубки раствор вводят в мышечную ткань, где роль АПК выполняют дендритные клетки. Попав в ядро дендритной клетки вакцина начинает экспрессировать, и происходит синтез белков-антигенов. С помощью микроинъекций ДНК также можно вводить подкожно, в тимус, печень, опухолевую ткань[16], однако именно в мышечной ткани наблюдается наиболее длительная (до года) экспрессия ДНК-вакцины[17].
Благодаря высокой концентрации клеток Лангерганса (подтип дендритных клеток), привлекательной мишенью для ДНК-вакцинации является кожа[18]. Для интрадермального (подкожного) введения используют массив из микроигл, длина которых несколько сотен микрон. Существуют различные варианты интрадермальной вакцинации. Простейший включает разрыхление массивом микроигл рогового слоя кожи (внешний слой кожи, обычно 10-20 мкм), чтобы увеличить её проницаемость для дальнейшего местного введения раствора ДНК. Более эффективно использование микроигл, покрытых сухой вакциной, которая растворяется уже под кожей[19].
Эффективность этого метода обычно низкая, поскольку сначала ДНК попадает в межклеточное пространство, а уже потом включается в клетки.
Электропорация[править | править код]

Электропорация — традиционный подход для доставки ДНК в бактериальные клетки и культуры клеток, который базируется на применении электрического импульса. Такой импульс создает поры в клеточной мембране, что способствует вхождению отрицательно заряженной ДНК. Этот способ был заимствован для доставки ДНК-вакцины в организм животных и человека и позволяет значительно повысить эффективность обычной инъекции. Прибор для электропорации имеет источник электрического тока и одноразовую сетку, которая состоит из шприца и игл-электродов. Шприц вводит вакцину в мышечную ткань, а электроды создают электрическое поле, которое облегчает вхождение ДНК в миоциты и дендритные клетки. На сегодня разработаны устройства, которые позволяют повысить эффективность вакцинации в 1000 раз по сравнению с обычной инъекцией. Электропорацию можно применять как для внутримышечного, так и для подкожного введения ДНК-вакцины. Недостатками являются незначительная болезненность в месте инъекции, потребность в специализированных устройствах[18]. Вместо электрического поля можно использовать магнитное. Такие устройства действуют по тому же принципу, однако в этом случае процедура является полностью безболезненной и менее повреждает клетки[20].
Действие электрического поля не только усиливает поглощение ДНК-вакцины клетками, но и стимулирует выработку иммунного ответа. Применение электропорации приводит к незначительному повреждению ткани — развивается локальный воспалительный процесс. Поврежденные клетки выделяют хемокины, поэтому к ним направляются макрофаги, лимфоциты и дендритные клетки. Увеличение концентрации иммунных клеток в месте введения вакцины повышает её эффективность[21].
Сонопорация[править | править код]
Сонопорация — метод переноса чужеродной ДНК в клетки с помощью ультразвука. Ультразвук увеличивает проницаемость клеточной мембраны, вследствие чего экзогенная ДНК легче проникает в клетку. Впервые сонопорация для переноса генов в клетку была применена в 1986 году[22]. Этот метод применяется для ввода молекул ДНК в клетки роговицы, мозга, костной ткани, почек, поджелудочной железы, эмбриональной ткани, скелетной и сердечной мышц. Относительно других методов сонопорация является малоисследованной, необходимо ещё немало усилий, чтобы повысить её эффективность, особенно на уровне целого организма[16].
Баллистическая трансфекция[править | править код]
Баллистическая трансфекция основывается на обстреливании (бомбардировке) органов и тканей микрочастицами тяжёлых металлов (золото, вольфрам) диаметром 1-3 мкм, покрытых молекулами ДНК. Введённая таким образом ДНК-вакцина экспрессируется в клетках-мишенях, а их продукты попадают в кровь. Для придания ускорения частицам используются похожее на стрелковое оружие устройство — генный пистолет, или генную пушку. Микрочастицы проходят через клеточные мембраны и переносят генетическую конструкцию непосредственно в ядро клетки. Глубина проникновения микрочастиц, как правило, невелика — до 1 мм, поэтому метод применяют преимущественно для трансфекции кожи или прилегающей хрящевой ткани. Особые условия обстрела позволяют микрочастицам проникать на глубину до 4-5 мм и переносить генные конструкции в волокна поперечно-полосатых мышц. Обычно клетки, находящиеся непосредственно по центру выстрела, погибают, в то время когда в зоне 0,6-1 см от центра находятся наиболее удачно протрансформированные клетки. Эту технологию называют также биобаллистикой или биолистикой[23].
Первый генный пистолет был создан группой учёных в период 1983 и 1986 годами с целью трансформации клеток растений. Это был пистолет, разработанный на основе устройства для автоматического забивания гвоздей. На вольфрамовый шар наносили ДНК с репортёрным (маркерным) геном и выстреливали им чашку Петри. Экспрессия репортёрного гена свидетельствовала об эффективности иммунизации. На сегодня для доставки ДНК используют частицы из золота или серебра, так как они не являются токсичными для клетки, в отличие от вольфрамовых[24].
Под действием высокого давления[править | править код]
В 1999 году были разработаны инъекционные приборы, которые способны вводить ДНК-вакцину без использования иглы[25]. Такие устройства работают благодаря силе Лоренца: небольшой мощный магнит создаёт значительное давление, приводит в действие поршень, который выбрасывает лекарственный препарат со скоростью звука[16]. Изменяя силу тока, можно выбирать глубину инъекции и дозировать лекарства. Процедура совершенно безболезненна и раньше использовалась для введения инсулина больным диабетом и при проведении масштабных вакцинаций. Существенным недостатком этого метода является то, что высокое давление теоретически может изменять структуру вводимых молекул[26]. Тем не менее, данную технологию ввода продолжают совершенствовать, и на сегодня разработаны приборы, которые могут доставить ДНК на глубину до 16 мм[27].
В составе живого бактериального вектора[править | править код]
Живые бактериальные векторы — это штаммы сальмонелла, шигеллы или листерия, которые несут мутации в генах биосинтеза или инвазии, что устраняет их патогенность и способность сохранять свою жизнеспособность в организме хозяина или окружающей среде. Взамен в геном бактерий встраивают нужные гены иммуногенных протеинов. Ослабленная бактерия вводится в организм пероральным путём (через рот, путём проглатывания) или интроназально (путём впрыска в носовое отверстие), поэтому этот способ вакцинации не требует никакого оборудования. Кроме того, такое введение стимулирует иммунный ответ слизистой оболочки, что важно, поскольку большинство патогенов попадают в организм через ротовое и назальное отверстия. Минуя желудок ослабленная бактерия попадает в тонкий кишечник. Далее бактерия проникает в Пейеровы бляшки — лимфоузлы кишечника. Оказавшись в середине Пейеровых бляшек, бактерии становятся мишенью для макрофагов и подвергаются фагоцитозу. В цитоплазме макрофагов происходит высвобождение бактерией ДНК-вакцины, после чего ДНК попадает в ядро, а бактерия обезвреживается иммунной системой[18][28].
Упаковка в липосомы[править | править код]


Липосома — шарообразное образование (около 100 нм в диаметре), состоящее из двойного липидного слоя. Липосомы имеют полость внутри, которая обычно заполнена растворителем, но может использоваться для доставки разнообразных веществ, в том числе и ДНК-вакцин. Их гидрофобная оболочка позволяет им сливаться с клеточными мембранами и переносить своё содержимое внутрь клетки. Использование липосом началось в 1965 г., и это стало мощным двигателем развития бионанотехнологий[29].
Перспективным способом прямого введения ДНК-конструкции в клетки-мишени является доставка генетической конструкции в составе катионных липосом, построенных из положительно заряженных липидов. Катионные липосомы с отрицательно заряженной молекулой ДНК образуют ДНК-липидный комплекс — липоплекс. Преимущества применения таких комплексов — способность нести большой объём информации, неинфекционность, кроме того, они просты и недороги в изготовлении[30]. В 2003 г. были созданы чрезвычайно малые — миллимикронные липосомы, покрытые полимером полиэтиленгликолем, которые способны проносить терапевтическую ДНК сквозь гемато-энцефалический барьер и доставлять её в нейроны головного мозга, что до этого было невозможным[31].
В составе полиплексов[править | править код]
Для введения в клетку ДНК-конструкций больших размеров (>10 т.п.н.) используют полиплексы — системы, состоящие из положительно заряженных полимеров (поликатионов) и отрицательно заряженных молекул ДНК. Размер таких комплексов составляет менее 100 нм, что, с одной стороны, не отдаёт их на переваривание макрофагам (поскольку они реагируют на частицы более 200 нм), а с другой стороны, они достаточно большие, чтобы не фильтроваться в почках[32].
Поликатионы конденсируют молекулу ДНК в комплексы, и таким образом обеспечивают её стабильность и защиту от действия нуклеаз. В качестве ДНК-связующих полимеров могут служить катионные белки, синтетические гомополимеры аминокислот (полилизины, полиаргинины), полисахарид хитозан, полиэтиленамин. Обычно в составе полиплексов поликатион находится в избытке, в результате чего данный комплекс является растворимым и положительно заряженным. Если к полиплексу пришить лиганд к определённому клеточному рецептору, то ДНК-вакцину можно направлять в конкретный тип клеток. Процесс доставки генетического материала в составе полиплексов включает два этапа: внеклеточный (путь от места введения к клеткам-мишеням) и внутриклеточный (взаимодействие с клетками-мишенями, эндоцитоз, выход из эндосом, доставка в ядро). Первым барьером, который необходимо преодолеть комплексу, является кровь и внеклеточный матрикс. Поэтому подбираются такие физико-химические параметры полиплекса, чтобы увеличить его стабильность, избежать нежелательных взаимодействий с белками крови и иммунной реакции, вызванной химической природой поликатиона. Попав в клетки-мишени, полиплекс адсорбируется на плазматической мембране, поглощается путём эндоцитоза, после чего он должен покинуть эндосому и диссоциировать на катионный полимер и молекулу ДНК. Свободная ДНК направляется в ядро, а катионый полимер покидает клетку и выводится из организма[33].
Метод
|
Преимущества
|
Недостатки
|
| Внутримышечные или подкожные инъекции |
|
|
| Электропорация |
|
|
| Генный пистолет |
|
|
| Введение за счёт высокого давления |
|
|
| Упаковка в липосомы |
|
|
Механизм развития иммунного ответа[править | править код]

Синтезированный в клетке антиген поддаётся процессингу, после чего происходит его представление иммунокомпетентным клеткам. Процессинг — это расщепление антигенного протеина на иммуногенные пептидные фрагменты. Представление означает подачу фрагмента антигена, соединённого с молекулами главного комплекса гистосовместимости (ГКГ) иммунокомпетентным клеткам. Различают два наиболее значимых класса этих молекул: ГКГ класса I (ГКГ-I) и ГКГ класса II (ГКГ-II). Для связывания с молекулами каждого класса антиген проходит подготовку в специализированных компартментах клетки. Эндогенные белки-антигены направляются на деградацию в протеасому, после чего представляются в комплексе с ГКГ-I на поверхности клетки. Здесь при встрече их распознают CD8+ T-клетки (Т-киллеры), которые реализуют цитотоксический иммунный ответ. Экзогенные белки расщепляются лизосомными протеазами, включаются в состав ГКГ-II и распознаются рецепторами CD4+ Т-клеток (Т-хелперов). Последние вызывают как клеточный, так и гуморальный ответ.
Пути процессинга антигена и формы иммунного ответа
| ||||
| Локализация антигена | Основной инструмент процессинга | Антигенпредставляющий комплекс | Клетки, которые распознают комплекс антиген-ГКГ |
Иммунный ответ |
|---|---|---|---|---|
| В клетке | Протеасома | ГКГ-I | Т-киллеры | Цитотоксический |
| Вне клетки | Лизосома | ГКГ-II | Т-хелперы | Клеточный, гуморальный |
Презентация антигена по пути ГКГ-I[править | править код]

ДНК-вакцинация предполагает эндогенный синтез антигена, поэтому этот путь является преобладающим. Процессинг антигена по пути ГКГ-I проходит в несколько этапов. Синтезированный в ядре белок транспортируется в цитоплазму, расщепляется протеасомой на короткие пептиды — эпитопы, которые специальными белками-транспортёрами антигенов (англ. ТАР, transporter associated with antigen processing) переносятся в эндоплазматический ретикулум (ЭПР). В ЭПР каждый эпитоп соединяется с молекулой ГКГ-I, после чего образованный комплекс направляется в аппарат Гольджи на гликозилирование. Оттуда комплекс в составе везикулы направляется к плазматической мембране. После слияния везикулы с плазмалеммой комплекс оказывается на поверхности клетки, где распознается рецепторами Т-киллеров, которые обладают цитотоксической активностью[34].
Презентация антигена по пути ГКГ-II[править | править код]
Основным источником пептидов, которые связываются с ГКГ-II, являются экзогенные белки, которые попали в клетку с помощью эндоцитоза. Однако показано, что некоторые внутриклеточные белки также могут быть представлены в комплексе с ГКГ-II[35]. При этом новосинтезированные белки после попадания в цитоплазму переносятся в лизосомы, где антиген расщепляется под действием кислых протеаз. После этого лизосома, которая содержит эпитопы, сливается с везикулой, которая несёт молекулу ГКГ-II. Внутри объединённой везикулы образуется комплекс эпитоп-ГКГ-II, который после слияния везикулы с плазмалеммой выносится на поверхность клетки. Здесь этот комплекс распознается рецепторами Т-хелперов, в результате чего происходит их активация. Это приводит к стимуляции как клеточного (активация Т-киллеров), так и гуморального иммунитета (активация В-лимфоцитов).
Традиционная вакцинация растворимыми белковыми антигенами нацелена на мобилизацию T-хелперов. Сравнительно низкий ответ Т-хелперов является одним из недостатков ДНК-вакцин. Также нынешнее поколение ДНК-вакцин не способно индуцировать продукцию высоких титров антител. Чтобы повысить активацию Т-хелперов, антиген нужно перенаправить на путь ГКГ-II. Для этого в ДНК-вакцину встраивают сигнал лизосомной локализации: синтезированный антиген будет направляться в лизосомы, а значит ступать на путь ГКГ-II[34].
Стратегии повышения эффективности ДНК-вакцины[править | править код]
Главный вопрос относительно будущего ДНК-вакцин касается повышения их эффективности. На сегодня проводится большое количество исследований, посвящённых оптимизации разработанных ДНК-вакцин. Поиск решения ведётся в двух направлениях: повышение экспрессии вакцины и увеличение иммуногенности закодированного антигена.
Оптимизация транскрипционных элементов[править | править код]
Важным компонентом ДНК-вакцины является промотор. Бактериальные промоторы не подходят для экспрессии антигена в клетках млекопитающих, поэтому вместо них использовали промоторы онкогенных вирусов. Сейчас, для повышения безопасности вакцин, их заменили на промоторы от неканцерогенных объектов, например, человеческого цитомегаловируса (CMV). Для большинства ДНК-вакцин этот промотор является оптимальным выбором: он характеризуется высокой экспрессией в широком диапазоне клеток. Для экспрессии гена в конкретных тканях, перспективным является использование промоторов, специфических для данного типа тканей. Например, использование промотора мышечной креатинкиназы при внутримышечном введении приводит к десятикратному увеличению синтеза антител и индукции Т-клеточного ответа, чем использование аналогичной ДНК-вакцины с CMV промотором. Также высокую эффективность в миоцитах показал промотор гена десмина, который кодирует один из белков цитоскелета. Для повышения экспрессии ДНК-вакцины в кератиноцитах (клетки эпителиальной ткани) используют промоторы гена металлотионеина (белок, который связывает тяжёлые металлы) или гена гидроксилазы витамина D3[13][36].
Уровень инициации транскрипции, как правило, повышается за счёт использования сильного промотора и энхансеров, а особенности терминации могут стать ограничивающим фактором. Эффективность полиаденилирования и процессинга первичного РНК-транскрипта меняется в зависимости от последовательности polyA-сигнала. То есть, последовательность полиаденилирования влияет на синтез антигена. Например, широко используемый polyA-сигнал вируса SV40 имеет меньшую эффективность, чем сигнал полиаденилирования гена β-глобина кролика или гена гормона роста быка[13].
Для эффективной трансляции мРНК млекопитающих должна иметь так называемую последовательность Козак. Вставка этой последовательности в ДНК-конструкцию может существенно увеличивать уровень синтеза антигена. Чтобы РНК-полимераза не проскочила стоп-кодон гена и не состоялся синтез удлинённого протеина, который не сможет потом получить правильную укладку, ген можно заканчивать двойным стоп-кодоном[36].
При конструировании ДНК-вакцины также пытаются оптимизировать её кодоны. Процедура оптимизации означает замену границ в последовательности гена таким образом, чтобы аминокислотная последовательность белка не менялась, но увеличивалась эффективность трансляции его мРНК. Причиной является то, что большинство аминокислот кодируются более чем одним кодоном. Каждый кодон имеет свою тРНК, и представленность различных тРНК в клетке неодинакова, причём она также варьирует в зависимости от вида организма. Кодоны подбирают таким образом, чтобы наличие нужной тРНК при синтезе антигена не стало лимитирующим фактором[13][37].
Оптимизация антигена[править | править код]
Хотя сила иммунного ответа коррелирует с уровнем экспрессии ДНК-вакцины, однако для каждого антигена существует определённое плато, после которого увеличение количества антигенного протеина не будет повышать продукцию антител. В то же время, достичь более сильной иммунной реакции можно за счёт оптимизации антигена[38]. Например, путём объединения антигена с лигандом до определённого рецептора антигенпредставляющей клетки (АПК). Таким лигандом может быть маркерный белок CD40, внеклеточный домен Fms-подобной тирозинкиназы-3 или антиген-4 Т-киллеров[39]. За счёт взаимодействия лиганд-рецептор повышается эффективность захвата антигенного протеина АПК.
Облегчение деградации антигена в протеасоме или лизосоме также будет стимулировать иммунную реакцию. Для усиления протелиотического расщепления антигена, в его последовательность встраивают сигнал убиквитинирования[34]. Использование кодирующих ДНК-вакцин вместо целого антигена, нескольких эпитопов различного происхождения, позволяет значительно расширить спектр иммунного ответа[38].
Для противоопухолевых ДНК-вакцин эффективной является комбинация «опухолевый антиген + вирусный или бактериальный антиген». Например, сочетание опухолевого антигена с эпитопом токсина столбняка значительно повышает активацию Т-киллеров против раковых клеток[40][41].
Включение адъювантов[править | править код]
При применении традиционных вакцин для повышения иммунного ответа к ним добавляют адъюванты. ДНК-вакцина имеет бактериальное происхождение, поэтому она сама является иммуностимулятором. Для усиления иммунного ответа в ДНК-вакцину встраивают гены адъюванта или применяют дополнительную плазмиду, которая кодирует иммуностимулирующие белки[42].
Иммуностимулирующее действие бактериальных CpG динуклеотидов[править | править код]
Функция плазмиды не ограничивается доставкой генов в клетки. Еще в 1893 году было обнаружено, что смесь бактериальных лизатов уменьшает прогрессирование раковых опухолей, однако лишь в 1983 установили, что иммуностимулирующие свойства лизата обусловлены молекулами ДНК бактерии[43]. В 1995 году показали, что стимуляция иммунитета вызвана CpG-мотивами бактериальной ДНК[44]. У бактерий, а также ДНК-вирусов, эти мотивы являются неметилированными. В организме человека и высших приматов, наоборот, цитозин в составе большинства CpG-динуклеотидов содержит метильную группу. Поэтому неметилированные CpG-мотивы воспринимаются человеческим организмом как патогенассоциированные молекулярные паттерны (ПАМП) (PAMP, pathogen-associated molecular patterns). ПАМП-соединения распознаются толл-подобными рецепторами, которых, в зависимости от типа лиганда, разделяют на несколько типов. Неметилированные CpG-мотивы распознаёт рецептор TLR-9, расположенный на мембранах эндоплазматического ретикулума В-лимфоцитов, дендритных клеток и натуральных киллеров. Связывание рецептора с неметилированными CpG-мотивами запускает каскад реакций, в результате которого индуцируется синтез провоспалительных цитокинов — интерферона-1 и IL-12[45].
Экспрессия цитокинов и других иммуномодуляторов[править | править код]
Для ДНК-вакцинации в качестве адъювантов чаще всего применяют гены цитокинов. Цитокины — это класс белковых молекул, регулирующих межклеточные и межсистемные взаимодействия в организме, в частности, функционирование иммунной системы. Все цитокины, а их известно более 30, могут модулировать иммунный ответ. Цитокины IL2, IL-12, интерферон γ, IL-15, IL-18 и IL-23 имеют стимулирующее влияние на Т-хелперы первого класса. К цитокинам, которые модулируют действие Т-хелперов второго класса, относятся: IL-4, IL-5, IL-6, IL-10 И IL-13. Тип цитокина подбирают согласно тому, какой тип иммунного ответа хотят усилить[46].
Различия иммунного ответа Т-хелперов 1 и 2 класса
| ||
| Отличия | Т-хелперы 1 | Т-хелперы 2 |
|---|---|---|
| Главная клетка-партнер | Макрофаг | В-клетка |
| Иммунный ответ | Клеточный. Усиливает действие макрофагов |
Гуморальный.
Стимулирует пролиферацию В-клеток |
Достичь повышения иммунного ответа можно с помощью хемокинов. Хемокины — это семейство цитокинов, которые способны вызвать хемотаксис чувствительных к ним клеток, в том числе и иммунных. В частности, рецепторы к хемокинам есть на антигенпредставляющих клетках хемокинов. Связывание хемокина со своим рецептором приводит к эндоцитозу комплекса антиген-хемокин внутрь АПК. Эта стратегия эффективно используется как при разработке противовирусных ДНК-вакцин[47], так и противоопухолевых[48].
Функцию адъюванта также может выполнять белок БТШ70 (белки теплового шока, англ. heat-shock proteins, HSP). Иммуностимулирующее действие БТШ70 основано на его способности выходить во внеклеточное пространство и связываться с рецепторами АПК. Механизм транспорта БТШ70 наружу пока окончательно не выяснен, но, скорее всего, существует несколько путей — экзоцитоз, секреция наружу или выход через канал[49]. Связывание БТШ70 со своим рецептором приводит к активации дендритных клеток, опосредует представление антигена и стимулирует продукцию хемокинов. Поскольку антиген слит с БТШ70, он также попадает во внеклеточное пространство, поэтому может представляться по пути ГКГ-II и активировать В-клетки. Во избежание аутоиммунных реакций для ДНК-вакцин используют бактериальный ген БТШ70[50].
Преимущества и недостатки ДНК-вакцин[править | править код]
Метод ДНК-вакцинации обладает рядом преимуществ, наиболее важным из которых является запуск как гуморального, так и клеточного иммунного ответа. Вакцины на основе ДНК обеспечивают долговременную экспрессию антигена и, соответственно, устойчивый иммунный ответ. К дополнительным факторам, способствующим развитию ДНК-иммунизации, относят простоту и низкую стоимость производства вакцины[51].
| Преимущества | Недостатки |
|---|---|
|
|
Применение ДНК-вакцин[править | править код]
ДНК-вакцины в ветеринарии[править | править код]
Все четыре одобренные Управлением США по контролю над продуктами питания и лекарственными препаратами вакцины созданы на основе плазмид. Для трёх из них рекомендован производителем способ введения — внутримышечно, для вакцины «ЛайфТайд» — внутримышечная инъекция, совмещённая с электропорацией. Если остальные вакцины направлены на активацию иммунитета, то для вакцины «ЛайфТайд» иммуностимулирующее действие является дополнительным. Продукт вакцины — соматолиберин, гормон, который стимулирует высвобождение гипофизом гормона роста и пролактина. Действие последних двух гормонов у свиней приводит к росту массы животных и увеличению численности выводка[52]. Вместе с тем, введение животным плазмиды, которая кодирует соматолиберин, стимулирует выработку Т-лимфоцитов, натуральных киллеров[53] следовательно, увеличивает иммунную сопротивляемость организма.
| Торговое название вакцины | Год лицензирования | Мишень | Животное | Продукт вакцины | Цель создания вакцины |
|---|---|---|---|---|---|
| «Вэст Найл-Инноватор» (США) Архивная копия от 4 октября 2013 на Wayback Machine | 2005 | Вирус лихорадки Западного Нила | Кони | Структурный белок вируса PreM-E | Защита против вируса |
| «Апекс-ай-эйч-эн» (Канада) Архивная копия от 4 октября 2013 на Wayback Machine | 2005 | Возбудитель инфекционного некроза гемопоэтической ткани (ИНГТ) | Рыбы семейства лососёвые | Вирусный гликопротеин | Повышение количества и качества продовольствия рыбы |
| «ЛайфТайд Эс-Даблъю 5» (Австралия) Архивная копия от 9 декабря 2012 на Wayback Machine | 2008 | Гормон роста | Свиньи и другой домашний скот | Соматолиберин свиньи | Увеличение выводка у свиноматок; значительно уменьшает перинатальную смертность и заболеваемость |
| «ОНСЕПТ» (США) | 2010 | Меланома | Собаки | Тирозиназа человека | Как альтернатива лучевой терапии и операционному вмешательству при лечении меланомы |
Перспективы ДНК-вакцинации[править | править код]

Противоопухолевые ДНК-вакцины[править | править код]
В то время как индукция клеточного и гуморального иммунного ответа убедительно продемонстрирована для чужеродных антигенов, ассоциированных с инфекционными заболеваниями, применение ДНК-вакцин для лечения рака до сих пор было менее успешным. Индукция эффективного противоопухолевого иммунитета является сложной задачей. Клинические исследования подтвердили общую безопасность и низкую токсичность противоопухолевых ДНК-вакцин, однако эффективность вызванного ими иммунного ответа оказалась слабой, а противоопухолевая активность, в некоторых случаях, вообще сомнительной[39].
ДНК-вакцины против вирусных и бактериальных возбудителей[править | править код]
Вакцины против вируса гепатита B[править | править код]

За последние тридцать лет для предупреждения возникновения инфекции вирусом гепатита B коммерческое использование получили семь вакцин. Все они основываются на использовании одного из вирусных оболочечных белков, называемого поверхностным антигеном, или HBsAg.
- Первая вакцина стала доступной в 1981—82 г, когда в Китае приступили к использованию вакцины, приготовленной из плазмы крови, полученной от доноров из числа больных, которые имели продолжительную инфекцию вирусного гепатита B. В этом же году она стала коммерчески доступна в США. Пик её применения пришёлся на 1982—88 гг. Вакцинацию проводили в виде курса из трёх прививок с временным интервалом. При постмаркетинговом наблюдении после введения такой вакцины отметили возникновение нескольких случаев побочных заболеваний центральной и периферической нервной системы, в том числе синдром Гийена-Барре, плекситы. В исследовании привитых вакциной лиц, проведённом через 15 лет, подтверждена высокая иммуногенность вакцины, приготовленной из плазмы крови.
- С 1987 г. на смену плазменной вакцине пришло следующее поколение вакцины против вируса гепатита B, в которой использована технология генной модификации рекомбинантной ДНК в клетках дрожжевого микроорганизма. Её иногда называют генно-инженерной вакциной. Синтезированный таким способом HBsAg выделяли из разрушаемых дрожжевых клеток. Ни один способ очистки не позволял избавляться от следов дрожжевых белков?[источник не указан 1489 дней] Новая технология отличалась высокой производительностью, позволила удешевить производство и уменьшить риск, происходящий из плазменной вакцины.
Для создания достаточно выраженной иммунной перестройки прививка состоит из трёх инъекций вакцины, причем вторую инъекцию делают через месяц после первой дозы, а третья инъекция — спустя шесть месяцев после первой дозы[55] После прививки поверхностный антиген вируса гепатита B можно обнаружить в сыворотке крови в течение нескольких дней; это называют вакцинной антигенемией. После этого в токе крови появляются антитела иммунной системы к HBsAg. Эти антитела известны как анти-HBsAg. С этого времени упомянутые антитела и память иммунной системы обеспечивают иммунитет к инфицированию гепатитом B.[56]
ДНК-вакцина против COVID-19[править | править код]
ДНК-вакцина INO-4800 производства Inovio Pharmaceuticals проходит в настоящее время клинические испытания I-II фаз[57][58]. Также клинические испытания проходят противоковидные ДНК-вакцины AG0301-COVID19, ZyCoV-D, GX-19, CORVax, а также ДНК-вакцины производства Symvivo и Entos Pharmaceuticals.
ДНК-вакцина против кариеса[править | править код]
Причиной кариеса является локальная смена pH вследствие брожения (гликолиза) углеводов, осуществляемого бактериями[59]. Учеными из Уханьского Института вирусологии (Китай) была разработана ДНК-вакцина, направленная против одного из возбудителей кариеса — Streptococcus mutans. Вакцина построена на основе плазмиды и кодирует два белка: поверхностный протеин St. mutans PAc и флагеллин, полученный из бактерии сальмонелла, который выполняет роль адъюванта[60]. На стадии доклинических исследований вакцину через нос вводили лабораторным грызунам, после чего у животных проверяли уровень иммуноглобулинов G в сыворотке крови и секреторных иммуноглобулинов А в слюне. После проведённых исследований учёные выяснили, что уровень иммунных белков и в крови и в слюне повышался, но, что важнее, при этом тормозился рост колоний Streptococcus mutans на зубной эмали. То есть, зубы вакцинированных животных были лучше защищены от кариеса.
ДНК-вакцины и лечение аутоиммунных заболеваний[править | править код]
ДНК-вакцина против диабета 1 типа[править | править код]
Сахарный диабет 1-го типа характеризуется потерей инсулин-продуцирующих бета-клеток, расположенных на островках Лангерганса поджелудочной железы. Главная причина потери бета-клеток — аутоиммунное поражение Т-киллерами[61]. Для того, чтобы защитить бета-клетки от гиперактивной функции иммунной системы, учёные из Стэнфордского (США)и Лейденского (Нидерланды) университетов, разработали ДНК-вакцину BHT-3021. Вакцина создана на основе плазмиды и кодирует предшественник инсулина — проинсулин. Это вакцина обратного действия: если обычные вакцины должны активировать иммунные реакции, то BHT-3021, наоборот, нейтрализует цитотоксическое действие Т-киллеров, направленную против островков Лангерганса.
В первой фазе клинических испытаний BHT-3021 показала свою эффективность на группе из 80 человек. Половина из них каждые семь дней в течение 12 недель получала внутримышечные инъекции BHT-3021, а вторая половина — плацебо. После окончания этого срока группа, которая получала вакцину, продемонстрировала повышение уровня С-пептидов в крови, что свидетельствует о восстановлении функции бета-клеток. Никаких серьезных побочных эффектов ни у кого из участников зафиксировано не было. Действие вакцины сохранялось в течение 2 месяцев[62].
Примечания[править | править код]
- ↑ Atanasiu P.; Cantarow A., Paschkis K. E. Production of neoplasms by fractions of mammalian neoplasms. (англ.) // Cancer Research : journal. — American Association for Cancer Research, 1950. — Vol. 10. — P. 775—782.
- ↑ Ito Y. Heat-resistance of the tumongenic nucleic acids of Shope papillomatous. (англ.) // Virology : journal. — 1961. — Vol. 12. — P. 596—601.
- ↑ Atanasiu P. Production de tureurs chez le Hamster par inoculation d’acide desoxyribonucleique extrait de cultures de tissus infectees par le virus du polyome. (фр.) // Acad. Sci. : magazine. — 1962. — Vol. 254. — P. 4228—4230.
- ↑ Orth G. Infectious and oncogenic effect of DNA extracted from cells infected with polyoma virus. (англ.) // PrOC. Soc. Exp. Biol. Med. : journal. — 1964. — Vol. 115. — P. 1090—1095.
- ↑ Wolff J. A.; Malone R. W., Williams P., Chong W. Direct gene transfer into mouse muscle in vivo. (англ.) // Science. — 1990. — Vol. 247, no. 4949. — P. 1465—1468. — PMID 1690918.
- ↑ Tang D.; Devit M.; Johnston S.A.; Others,. Genetic immunization is a simple method for eliciting an immune response (англ.) // Nature : journal. — 1992. — Vol. 356, no. 6365. — P. 152—154. — doi:10.1038/356152a0. — PMID 1545867.
- ↑ Ulmer J. B.; Donnelly J., Parker S. E., Rhodes, Others,. Heterologous protection against influenza by injection of DNA encoding a viral protein. (англ.) // Science : journal. — 1993. — Vol. 259. — P. 1745—1749.
- ↑ Wang B; Ugen K. E., Srikantan V., Agadjanyan M. G.; Others,. Gene inoculation generates immune responses against human immunodeficiency virus type 1 (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 90, no. 9. — P. 4156—4160. — doi:10.1073/pnas.90.9.4156.
- ↑ Conry R. M.; LoBuglio, A. F., Loechel, F., Moore, S. E.; Others,. A carcinoembryonic antigen polynucleotide vaccine for human clinical use (каталан.) // Cancer gene therapy. — 1995. — Vol. 2, num. 1. — P. 33—38.
- ↑ Chattergoon M; Boyer J., Weiner DB. Genetic immunization: a new era in vaccines and immune therapeutics (англ.) // The FASEB Journal : journal. — Federation of American Societies for Experimental Biology, 1997. — Vol. 11, no. 10. — P. 753—763. — PMID 9271360.
- ↑ Ferraro B.; Matthew P. Morrow, Natalie A. Hutnick; Others. Clinical Applications of DNA Vaccines: Current Progress (англ.) // Clin Infect Dis. : journal. — 2011. — Vol. 53, no. 3. — P. 296—302. — doi:10.1093/cid/cir334.
- ↑ Anderson R. J.; Schneider J. Plasmid DNA and viral vector-based vaccines for the treatment of cancer (англ.) // Vaccine : journal. — Elsevier, 2007. — Vol. 25. — P. 24—34. — doi:10.1016/j.vaccine.2007.05.030..
- ↑ 1 2 3 4 Garmory H. S.; Brown K. A., Titball R. W. DNA vaccines: improving expression of antigens (англ.) // Genetic Vaccines and Therapy. — 2003. — Vol. 1, no. 1. — P. 2. — doi:10.1186/1479-0556-1-2.
- ↑ Супотницкий М. В. ДНК-иммунизация в профилактике инфекционных заболеваний сельскохозяйственных животных – 1998. – Т. 5. – С. 18-24. // Ветеринария : журнал. — 1998. — Т. 5. — С. 18—24. Архивировано 7 декабря 2013 года.
- ↑ Tang D.; Ottensmeier C. H., Stevenson F. K. DNA vaccines: precision tools for activating effective immunity against cancer (англ.) // Nature Reviews Cancer : journal. — 2008. — Vol. 8, no. 2. — P. 108—120. — doi:10.1038/nrc2326.
- ↑ 1 2 3 4 Kamimura K.; Suda T., Zhang G., Liu D. Advances in gene delivery systems (англ.) // Pharmaceutical medicine. — 2011. — Vol. 5. — P. 293—306.
- ↑ Wolff J. A.; Ludtke J. J., Acsadi G., Williams P., Jani A. Long-term persistence of plasmid DNA and foreign gene expression in mouse muscle (англ.) // Hum. Mol. Gen : journal. — 1992. — Vol. 1. — P. 363—369.
- ↑ 1 2 3 4 Cranenburgh R. DNA Vaccine Delivery (англ.) // BioPharm International Supplements. — 2011. — Vol. 24, no. 10. — P. 12—18. Архивировано 21 сентября 2013 года.
- ↑ Chen X; Kask A. S., Crichton M. L., Others,. Improved DNA vaccination by skin-targeted delivery using dry-coated densely-packed microprojection arrays (англ.) // J Control Release : journal. — 2010. — Vol. 148, no. 3. — P. 327—333. — doi:10.1016/j.jconrel.2010.09.001.
- ↑ Chen C.; Evans J. A., Robinson M. P., Others,. Genetic immunization is a simple method for eliciting an immune response (англ.) // Physics in Medicine and Biology : journal. — 2010. — Vol. 55, no. 4. — P. 1219—1223. — doi:10.1088/0031-9155/55/4/021.
- ↑ Kjeken R.; Devit M.; Johnston S.A.; Others,. Recruitment of antigen-presenting cells to the site of inoculation and augmentation of human immunodeficiency virus type 1 DNA vaccine immunogenicity by in vivo electroporation (англ.) // Journal of Virology : journal. — 2008. — Vol. 82, no. 11. — P. 5643—5649. — doi:10.1128/JVI.02564-07. — PMID 1545867.
- ↑ Fechheimer M.; Boylan J. F., Parker S., Others,. Transfection of mammalian cells with plasmid DNA by scrape loading and sonication loading (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1987. — Vol. 84, no. 23. — P. 8463—8467. — PMID 2446324.
- ↑ Абрамова, З.И. (2008), Введение в генетическую инженерию (PDF), Казань, pp. 110—116
{{citation}}: Википедия:Обслуживание CS1 (отсутствует издатель) (ссылка) Источник. Дата обращения: 21 декабря 2013. Архивировано 2 октября 2013 года. - ↑ Klein, TM et al (1987) High-velocity microprojectiles for delivering nucleic acids into living cells Архивная копия от 27 сентября 2013 на Wayback Machine. Nature 327:70-73
- ↑ Liu F; Song Y., Liu D. Hydrodynamics-based transfection in animals by systemic administration of plasmid DNA (англ.) // Gene Ther. : journal. — 1999. — Vol. 6, no. 7. — P. 1258—1266. — doi:10.1038/356152a0. — PMID 10455434.
- ↑ Kumaragurubaran K.; Kaliaperumal K. DNA vaccine: the miniature miracle (англ.) // Vet World. — 2013. — Vol. 6, no. 4. — P. 228—232. — doi:10.5455/vetworld.2013.228-232.
- ↑ Taberner A.; Devit M., Hogan C. N., Hunter I. W. Needle-free jet injection using real-time controlled linear Lorentz-force actuators (англ.) // Medical engineering & physics : journal. — 2012. — Vol. 34, no. 9. — P. 1228—1235. — doi:10.1016/j.medengphy.2011.12.010.
- ↑ Gentschev I.,; Dietrich G., Spreng S., Pilgrim S., Stritzker J., Kolb-Mäurer A., Goebel W. Delivery of protein antigens and DNA by attenuated intracellular bacteria (англ.) // Journal of Medical Microbiology : journal. — Microbiology Society, 2002. — Vol. 291. — P. 577—582. — PMID 11890559.
- ↑ Balazs D. A.; Godbey W. T. Liposomes for Use in Gene Delivery (англ.) // Journal of Drug Delivery. — 2011. — Vol. 2011. — P. 12. — doi:10.1155/2011/326497.
- ↑ Великий Николай Николаевич, Генная терапия: достижения, перспективы Источник. Дата обращения: 21 декабря 2013. Архивировано из оригинала 21 сентября 2013 года.
- ↑ Ananthaswamy, Anil. Undercover genes slip into the brain. New Scientist (20 марта 2003). Дата обращения: 17 августа 2010. Архивировано 8 января 2014 года.
- ↑ Moghimi S. M.; Hunter A. C., Murray J. C. Nanomedicine: current status and future prospects (англ.) // The FASEB Journal : journal. — Federation of American Societies for Experimental Biology, 2005. — Vol. 19. — P. 311—330. — doi:10.1096/fj.04-2747rev.
- ↑ Дурыманов Михаил, Доставка генов в клетку Источник. Дата обращения: 21 декабря 2013. Архивировано 21 сентября 2013 года.
- ↑ 1 2 3 Стародубова Е. С.; Исагулянц М. Г., Карпов В. Л. Регуляция процессинга иммуногена: сигнальные последовательности и их использование для создания нового поколения ДНК-вакцин // ACTA NATURAE : журнал. — 2010. — Т. 2, № 1. — С. 59—65.
- ↑ Klionsky D.; Emr. S. Autophagy as a regulated pathway of cellular degradation (англ.) // Science : journal. — 2000. — Vol. 290. — P. 1717—1721. — doi:10.1126/science.290.5497.1717.
- ↑ 1 2 Tang D.; Weiner D. B. DNA vaccines: ready for prime time? (англ.) // Nature Reviews Genetics : journal. — 2008. — Vol. 9, no. 10. — P. 776—788. — doi:10.1038/nrg2432.
- ↑ Williams J.A. Vector Design for Improved DNA Vaccine Efficacy, Safety and Production (англ.) // Vaccines : journal. — 2013. — Vol. 1, no. 3. — P. 225—249. — doi:10.3390/vaccines1030225. — PMID 1545867.
- ↑ 1 2 Moreno S; Timon M. DNA vaccination: an immunological perspective (англ.) // Immunologia. — 2004. — Vol. 123. — P. 41—55. Архивировано 21 сентября 2013 года.
- ↑ 1 2 Shaw D. R.; Strong T. V. DNA vaccines for cancer (англ.) // Frontiers in Bioscience : journal. — Frontiers in Bioscience, 2006. — Vol. 11. — P. 1189—1198. Архивировано 21 сентября 2013 года.
- ↑ Rice J.; Dossett M. L., Ohlen C., Buchan S. L., Others. DNA fusion gene vaccination mobilizes effective anti-leukemic cytotoxic T lymphocytes from a tolerized repertoire (англ.) // European Journal of Immunology : journal. — 2008. — Vol. 38, no. 8. — P. 2118—2129. — doi:10.1002/eji.200838213.
- ↑ Chudley L.; McCann K., Mander A., Tjelle T., Others. DNA fusion-gene vaccination in patients with prostate cancer induces high-frequency CD8+ T-cell responses and increases PSA doubling time (англ.) // Cancer Immunol Immunother : journal. — 2012. — Vol. 61. — P. 2161—2170. — doi:10.1002/eji.200838213.
- ↑ Ulmer J. B.; Wahren B., Liu M. A. Dna vaccines: recent technological and clinical advanced (англ.) // Discovery Medicine, 6(33):109-112, 2006. — 2006. — Vol. 6, no. 33. — P. 109—112. Архивировано 26 сентября 2013 года.
- ↑ Coley, W. B. The treatment of malignant tumors by repeated inoculations of erysipelas. With a report of ten original cases. 1893 (англ.) // Clinical orthopaedics and related research : journal. — 1991. — No. 262. — P. 3—11. — PMID 1984929.
- ↑ Krieg, AM; Yi, AK; Matson, S; Waldschmidt, TJ; Bishop, GA; Teasdale, R; Koretzky, GA; Klinman, D. M. CpG motifs in bacterial DNA trigger direct B-cell activation (англ.) // Nature : journal. — 1995. — Vol. 374, no. 6522. — P. 546—549. — doi:10.1038/374546a0. — PMID 7700380.
- ↑ Entrez Gene: TLR9 toll-like receptor 9. Архивировано 18 сентября 2019 года.
- ↑ Thalhamer, Josef; Weiss, Richard; Scheiblhofer, Sandra (2010), Gene Vaccines, Berlin: Springer, pp. 198—203, doi:10.1007/978-3-7091-0439-2
- ↑ Hong Qin; Pramod N. Nehete, Hong He, Others,. Prime-Boost Vaccination Using Chemokine-Fused gp120 DNA and HIV Envelope Peptides Activates Both Immediate and Long-Term Memory Cellular Responses in Rhesus Macaques (англ.) // Journal of Biomedicine and Biotechnology : journal. — 2010. — Vol. 2010. — P. 152—154. — doi:10.1155/2010/860160.
- ↑ Igoucheva, O; Grazzini M., Pidich A., Others,. Immunotargeting and eradication of orthotopic melanoma using a chemokine-enhanced DNA vaccine (англ.) // Gene Therapy : journal. — 2013. — Vol. 20, no. 9. — P. 938—949. — doi:10.1038/gt.2013.17.
- ↑ Евдонин А. Л.; Медведева Н. Д. Внеклеточный белок теплового шока 70 и его функции (англ.) // Цитология. — 2009. — Vol. 51, no. 2. — P. 130—137.
- ↑ Ebrahimi S. M.; Tebianian M. Immunotargeting and eradication of orthotopic melanoma using a chemokine-enhanced DNA vaccine (англ.) // World Applied Sciences Journal : journal. — 2013. — Vol. 14, no. 10. — P. 1569—1575. Архивировано 29 сентября 2013 года.
- ↑ doi: 10.1111/j.1600-065X.2010.00980.x
- ↑ Draghia-Akli R.; Ellis K. M., Hill L.-A., P.; Others,. High-efficiency growth hormone releasing hormone plasmid vector administration into skeletal muscle mediated by electroporation in pigs (англ.) // The FASEB Journal : journal. — Federation of American Societies for Experimental Biology, 2003. — doi:10.1096/fj.02-0671fje.
- ↑ Brown P. A.; Davis W. C., Draghia-Akli R. Immune-enhancing effects of growth hormone-releasing hormone delivered by plasmid injection and electroporation. (англ.) // Mol Ther. : journal. — 1992. — Vol. 10, no. 4. — P. 644—651. — doi:10.1038/356152a0. — PMID 15451448.
- ↑ Ferraro B.; Morrow, M. P., Hutnick, N. A. Clinical applications of DNA vaccines: current progress (англ.) // Clinical infectious diseases : journal. — 2011. — Vol. 53, no. 3. — P. 296—303. — doi:10.1093/cid/cir334.
- ↑ Hepatitis B Vaccine Information from Hepatitis B Foundation: Архивная копия от 28 июня 2011 на Wayback Machine
- ↑ Centers for Disease Control, USA (December 8, 2006). «Hepatitis B Vaccine: Fact Sheet». Дата обращения: 2 августа 2021. Архивировано 23 июня 2008 года.
- ↑ Tung Thanh Le, Zacharias Andreadakis, Arun Kumar, Raúl Gómez Román, Stig Tollefsen. The COVID-19 vaccine development landscape (англ.) // Nature Reviews Drug Discovery. — 2020-04-09. — Vol. 19, iss. 5. — P. 305–306. — doi:10.1038/d41573-020-00073-5. Архивировано 10 мая 2020 года.
- ↑ IVI, INOVIO, and KNIH to partner with CEPI in a Phase I/II clinical trial of INOVIO’s COVID-19 DNA vaccine in South Korea (англ.). nternational Vaccine Institute. Дата обращения: 29 декабря 2020. Архивировано 24 сентября 2020 года.
- ↑ Introduction To Dental Plaque (англ.). School of Dentistry. University of Leeds. Дата обращения: 17 марта 2009. Архивировано 9 августа 2007 года.
- ↑ Shi W. et al. Flagellin enhances saliva IgA response and protection of anti-caries DNA vaccine (англ.) // Journal of dental research : journal. — 2012. — Vol. 91, no. 3. — P. 249—254.
- ↑ Type 1 Diabetes Mellitus. Дата обращения: 4 августа 2008. Архивировано 21 июля 2013 года.
- ↑ Roep B. O.; Solvason, N., Gottlieb, P. A., Abreu, J. R. Plasmid encoded proinsulin preserves C-peptide while specifically reducing proinsulin-specific CD8 T cells in type 1 diabetes (англ.) // Sci Transl Med. : journal. — 2013. — Vol. 5, no. 191. — P. 191—182. — doi:10.1126/scitranslmed.3006103.
Литература[править | править код]
- Логунов Д. Ю., Народицкий Б. С., Гинцбург А. Л. Молекулярно-генетические технологии защиты от патогенов // «Ремедиум» : журнал. — М.: ООО «Группа Ремедиум», 2007. — № 3. — ISSN 1561-5936. Архивировано 29 октября 2013 года.
- Максютов Р. А., Щелкунов С. Н. Сравнение протективных свойств противооспенной ДНК-вакцины на основе гена A30L вируса натуральной оспы и его варианта с измененным кодоновым составом // Молекулярная генетика, микробиология и вирусология : научный журнал. — М.: Институт молекулярной генетики РАН, Издательство «Медицина», 2011. — № 2. — ISSN 0208-0613.
- Моисеенко В. М. Возможности вакцинотерапии меланомы кожи // «Практическая онкология» : научный журнал. — СПб.: ООО «Центр ТОММ», 2001. — Декабрь (№ 4 (8)). — С. 58–64.
- Петров Р. В., Хаитов Р. М. Иммуногены и вакцины нового поколения. Руководство. — М.: ГЭОТАР-Медиа, 2011. — 608 с. — (Библиотека врача-специалиста). — ISBN 978-5-9704-1868-0.
- Сергеев В. А., Непоклонов Е. А., Алипер Т. И. ДНК-вакцины // Вирусы и вирусные вакцины. — М.: «Библионика», 2007. — ISBN 5-98685-012-2.
- Ferraro B.; Morrow, M. P., Hutnick, N. A. Clinical applications of DNA vaccines: current progress (англ.) // Clinical infectious diseases : journal. — 2011. — Vol. 53, no. 3. — P. 296—303. — doi:10.1093/cid/cir334.
- Tang D.; Weiner D. B. DNA vaccines: ready for prime time? (англ.) // Nature Reviews Genetics : journal. — 2008. — Vol. 9, no. 10. — P. 776—788. — doi:10.1038/nrg2432.
- Irsa Mateen; Saba Irshad. A Review on DNA Vaccines (неопр.) // Journal of Health Science. — 2011. — Т. 1, № 1. — С. 1—7. — doi:10.5923/j.health.20110101.01.
- Hooper J. W., Thompson E., Wilhelmsen C., et al. Smallpox DNA vaccine protects nonhuman primates against lethal monkeypox (англ.) // J. Virol. : journal. — 2004. — May (vol. 78, no. 9). — P. 33—43. — doi:10.1128/JVI.78.9.4433-4443.2004.
Ссылки[править | править код]
- ДНК-вакцины: создание и перспективы. (рус.) (недоступная ссылка) Архивировано 19 февраля 2010 года.
- Видео: Клонирование ДНК-вакцины в бактериальной клетке. Архивная копия от 27 сентября 2016 на Wayback Machine (англ.)
- Видео: ДНК-вакцина против ВИЧ. Архивная копия от 26 декабря 2013 на Wayback Machine (рус.)